

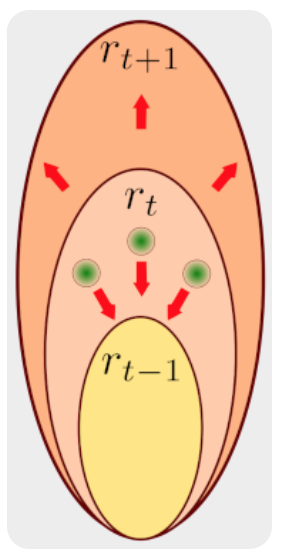
The front velocity, is related to particles velocity through local density variations,

Particles velocity is determined according to the geometry of the outlet. In flat-bottomed hoppers, this is determined using kinematic model, obtained by solving a diffusion type equation.

In conical hoppers particle velocity is assumed radial as determined through the plasticity model, the flow is

The solution of this set of equations leads to the evolution of the dilated and draw zones which are useful for the optimization of the mine operation.
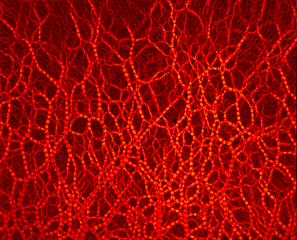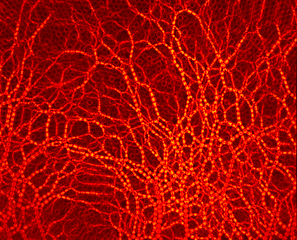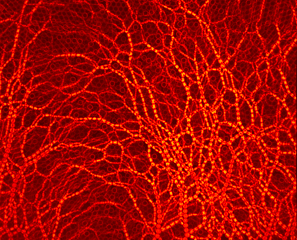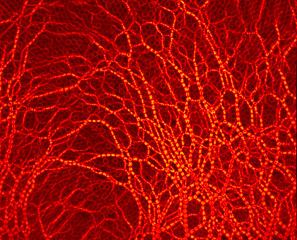
Photoelastic methodology reveals cavity formation induced by the depletion of stresses due to the formation of arches in a single draw-point operation.
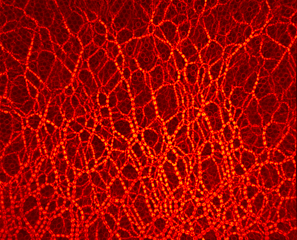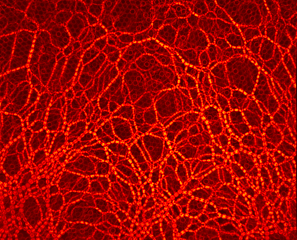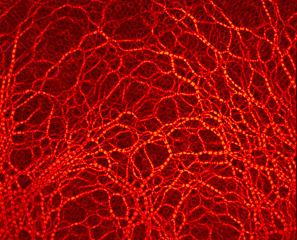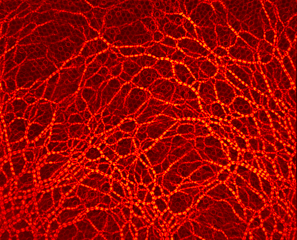
Photoelasticity evidence redistribution of contact forces during particles extraction from two neighboring draw-points simultaneously.